
 敬请关注公众号
敬请关注公众号

蛋白质翻译水平调控一直是植物生长发育重要调控方式,由于多年来研究手段缺失、核糖体组成成分复杂性高等原因,对于翻译调控方式的研究进展相较于表观水平调控、转录水平调控、翻译后水平调控更为缓慢。近年来对翻译水平调控,包括5‘mRNA加工、3’端加工、mRNA剪接、tRNA修饰、蛋白重翻译等众多过程已有相关研究。在众多的蛋白翻译调控过程中,真核生物延伸因子3 H亚基(eIF3h)在蛋白质重翻译过程起着举重轻重的地位。作为蛋白翻译调控的“小刹车”,它是如何通过影响mRNA的非翻译开放阅读框(uORF)和开放阅读框(ORF)的核糖体复合体招募,进而影响蛋白质翻译过程?这些问题值得我们深入探讨和研究。
位点信息
Locus: AT1G10840
Name:TIF3H1, TRANSLATION INITIATION FACTOR 3 SUBUNIT H1
基因功能
真核生物延伸因子(eIFs)是一类在蛋白质核糖体形成与行使功能上有着重要作用的延伸因子,这些延伸因子高度保守,而且对无论动植物生长发育扮演重要的角色。eIF3是这些延伸因子中组成成分最多,最为复杂的复合体。它主要能够结合40S抑制60S过早形成核糖体;eIF3可以促进43S PIC复合体的生成;eIF3参与核糖体对mRNA招募和扫描寻找ORF;eIF3还可以作为信号通路调控的节点平台整合不同信号,并传递到核糖体翻译过程,对蛋白质翻译起始微调控作用。在拟南芥中,eIF3含有11个不同的亚基。eIF3h亚基主要参与以ARF为主的uORF重翻译过程,并且受到TOR和S6K1磷酸化影响。水稻中报道eIF3h与脚手架蛋白RACK1和tRNAHis鸟苷转移酶相互作用调控植物高温条件下热适应性。
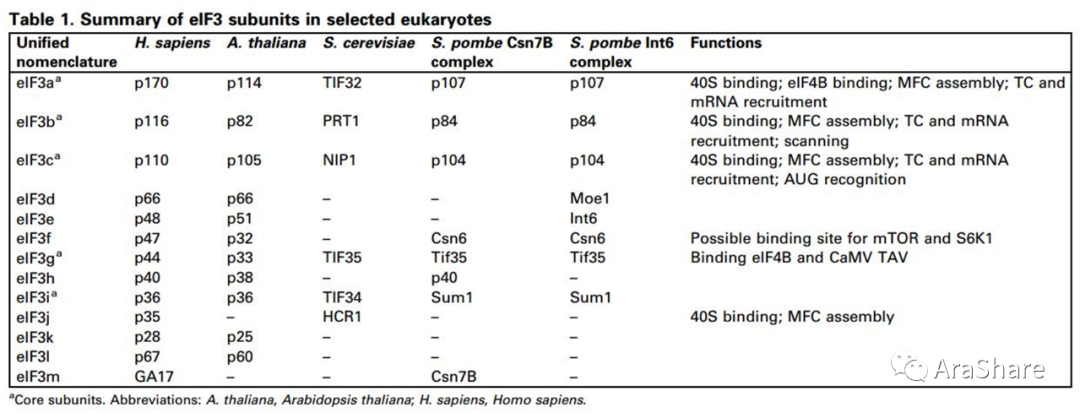
图1 真核生物eIF3亚基分布及其功能[1]
突变体信息
国内拟南芥共享中心AraShare有2个eif3h突变体,分别是SN17813(SALK_094155C)和SN18367(SALK_127106C)。突变体表现出多种生长劣势,育性和花器官发育均受到明显影响。对外源糖和ABA高度敏感。种子结实率低。

图2 eif3h突变体与野生型表型[2]
基因表达谱
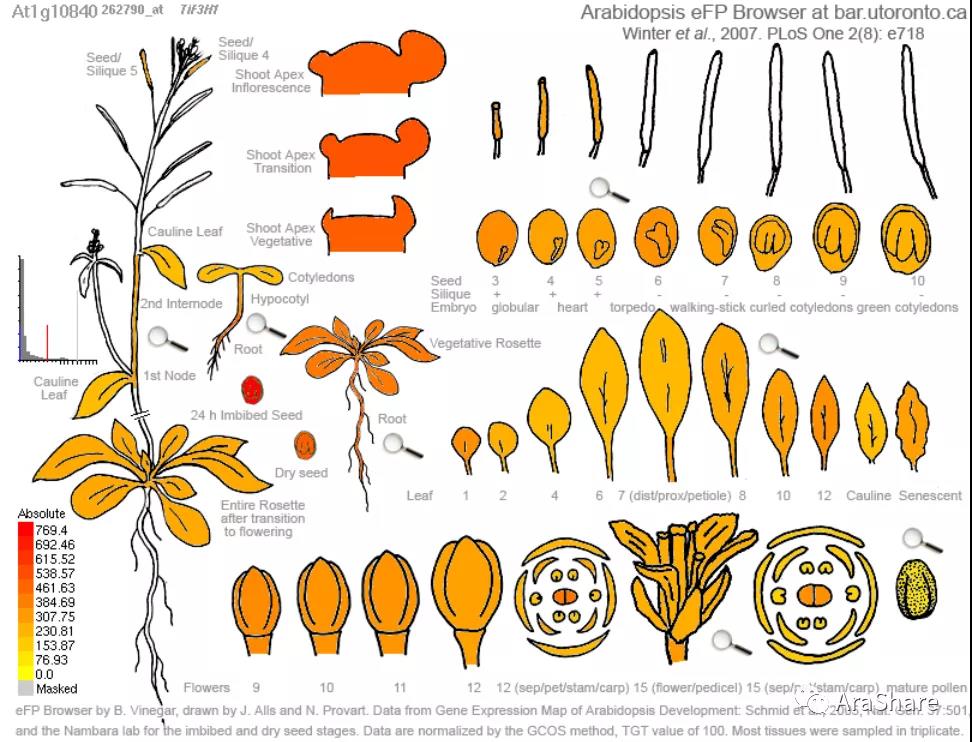
图3 拟南芥eIF3h基因表达谱(Source:eFP Browser http://www.bar.utoronto.ca/efp/cgi-bin/efpWeb.cgi)
研究概况
2004年,美国田纳西大学Albrecht G. von Arnim课题组在the Plant Cell发表题为“Translational Regulation via 59 mRNA Leader Sequences Revealed by Mutational Analysis of the Arabidopsis Translation Initiation Factor Subunit eIF3h”。这个基因是eIF3在植物中非必需亚基,因此不是一般蛋白质翻译所必须的。然而,他们发现在一些bZIP转录因子翻译水平受到明显调控,eIF3h与COP9 signalosome (CSN)蛋白相互作用。表明eIF3h参与了特定的mRNA翻译调控过程。
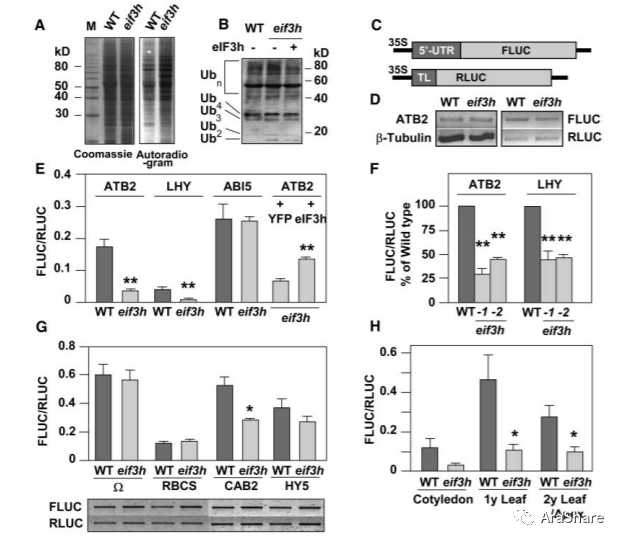
图4 eIF3h参与特定mRNA翻译调控过程[2]
2007年,Albrecht G von Arnim在Genome Biology发表了“On the functions of the h subunit of eukaryotic initiation factor 3 in late stages of translation initiation”。他们利用对野生型和eif3h突变体蛋白质组、微阵列等相关方法确认了eIF3h参与了含有uORF结构的mRNA蛋白翻译的重要调控过程[3]。2年后,他们在RNA杂志发表了“The h subunit of eIF3 promotes reinitiation competence during translation of mRNAs harboring upstream open reading frames”,利用荧光素酶报告系统互换不同的启动子序列,在野生型和eif3h突变体观察荧光素酶的两来确定翻译效率,这种方法最终确认eIf3h对重翻译过程的识别作用不大,而主要作用是促进重翻译过程再起始。
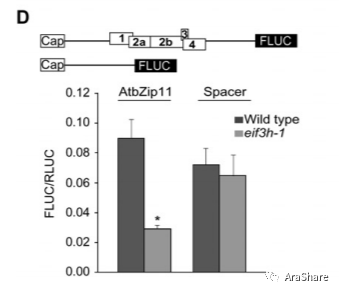
图5 体外荧光素酶实验证明AtZip11翻译效率在eif3h突变体受到显著抑制[4]
2013年,法国Université de Strasbourg的Lyubov A Ryabova课题组在The EMBO Journal发表“TOR and S6K1 promote translation reinitiation of uORF-containing mRNAs via phosphorylation of eIF3h”。作者用多种生化手段确认mTOR-S6K1-eIF3h的磷酸化信号调控网络,并且将生长素信号和eIF3h联系起来。同时也揭示了mTOR信号在蛋白质重翻译过程起着重要作用[5]。2014年,PLoS One 刊登了“Translational Control of Arabidopsis Meristem Stability and Organogenesis by the Eukaryotic Translation Factor eIF3h”的研究论文,把eIF3h调控的下游对象拓展至WUSCHEL (WUS) 和 CLAVATA (CLV1/3)分生组织的重要调控系统,包括ASYMMETRIC LEAVES 1 (AS1)在内的含有uORF的mRNA都受到eIF3h的调控。部分解释eif3h突变体在发育上的表型[6]。
其他物种研究情况简介
2019年,中科院上海卓越创新中心林鸿宣课题组在Molecular Plant发表了题为“Translational Regulation of Plant Response to High Temperature by a Dual-Function tRNAHis Guanylyltransferase in Rice”的研究论文。这篇研究论文发现AET1-eIF3h-RACK1三者相互作用,共同调控含有uORF结构的ARF基因蛋白翻译过程,可能也涉及蛋白重翻译过程[7]。
展望
eIF3h参与的蛋白重翻译调控过程的神秘面纱在过去近15年时间不断被揭开,但仍然留下了许多未解的问题:eIF3h具体是如何重新招募蛋白到ORF?在不同的生物胁迫或者细胞微环境下,eIF3h是如何选择特定mRNA?是否存在eIF3h调控的motif识别pattern?是否还有其它成员也参与到蛋白重翻译过程?除了磷酸化eIF3h,eIF3h还能受到哪些蛋白翻译后修饰影响?是否可能寻找到eIF3h的转录水平调控元件?当我们真正理解了蛋白重翻译过程,才能对这一翻译水平的“小刹车“有更深的理解。近年来上述一些作者也写了相关蛋白重翻译过程的综述,读者可以更深理解eIF3h在这其中扮演的角色[8–10]。
参考文献
[1] Hinnebusch, A.G. eIF3: a versatile scaffold for translation initiation complexes. Trends Biochem. Sci. 2006, 31, 553–562.
[2] Kim, T.-H.; Kim, B.-H.; Yahalom, A.; Chamovitz, D.A.; von Arnim, A.G. Translational Regulation via 5′ mRNA Leader Sequences Revealed by Mutational Analysis of the Arabidopsis Translation Initiation Factor Subunit eIF3h. Plant Cell 2004, 16, 3341–3356.
[3] Kim, B.; Cai, X.; Vaughn, J.N.; von Arnim, A.G. On the functions of the h subunit of eukaryotic initiation factor 3 in late stages of translation initiation. Genome Biol. 2007, 8, R60.
[4] Roy, B.; Vaughn, J.N.; Kim, B.H.; Zhou, F.; Gilchrist, M.A.; Von Arnim, A.G. The h subunit of eIF3 promotes reinitiation competence during translation of mRNAs harboring upstream open reading frames. Rna 2010, 16, 748–761.
[5] Schepetilnikov, M.; Dimitrova, M.; Mancera-Martínez, E.; Geldreich, A.; Keller, M.; Ryabova, L.A. TOR and S6K1 promote translation reinitiation of uORF-containing mRNAs via phosphorylation of eIF3h. EMBO J. 2013, 32, 1087–1102.
[6] Zhou, F.; Roy, B.; Dunlap, J.R.; Enganti, R.; von Arnim, A.G. Translational Control of Arabidopsis Meristem Stability and Organogenesis by the Eukaryotic Translation Factor eIF3h. PLoS One 2014, 9, e95396.
[7] Chen, K.; Guo, T.; Li, X.M.; Zhang, Y.M.; Yang, Y.B.; Ye, W.W.; Dong, N.Q.; Shi, C.L.; Kan, Y.; Xiang, Y.H.; et al. Translational Regulation of Plant Response to High Temperature by a Dual-Function tRNAHis Guanylyltransferase in Rice. Mol. Plant 2019, 12, 1123–1142.
[8] Schepetilnikov, M.; Ryabova, L.A. Auxin signaling in regulation of plant translation reinitiation. Front. Plant Sci. 2017, 8, 1–15.
[9] Merchante, C.; Stepanova, A.N.; Alonso, J.M. Translation regulation in plants: an interesting past, an exciting present and a promising future.Plant J. 2017, 90, 628–653.
[10] Urquidi Camacho, R.A.; Lokdarshi, A.; von Arnim, A.G. Translational gene regulation in plants: A green new deal. Wiley Interdiscip. Rev. RNA2020, 1–40.
本推文基于科学文献和网络资讯写作而成,未经领域内专家审阅;限于笔者学识水平,难免存在错漏,仅供参考!